
�������: 1-15 ���鵽������ҽѧ ATP����ؼ�¼46�� . ��ѯʱ��(0.187 ��)

2021��11��8�գ�������ѧ����ҽѧԺ���ֿ�������Nature Communications��־���߷�������ΪStructure and activation mechanism of the hexameric plasma membrane H+-ATPase���о����ģ�https://doi.org/10.1038/s41467-021-26782-y�������о������������ƣ�3.2 ���ͼ�...
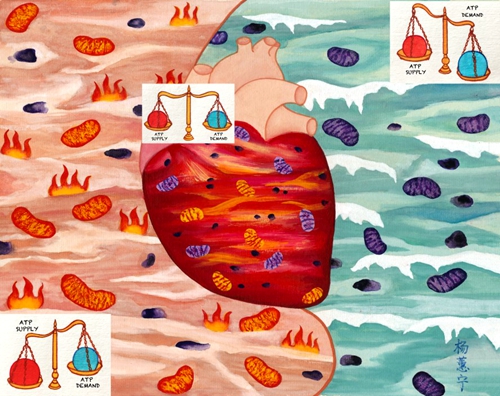
������ѧ����ҽѧ�о������Ի��������ʾ�������ŵ�������ATP��̬ƽ�⣨ͼ��
������ѧ����ҽѧ�о��� ���Ի������� �������� ����ATP ��̬ƽ��
2017/7/18
ATP��ϸ�����������ң�Ҳ�������������̵���Ҫ�ź����ʡ����������£�ϸ����ATPŨ��ͨ��ά����ʮ���ȶ���ˮƽ�������Ƕ��ڸߺ��ܵ����࣬�����˶�ʱ����ATP�IJ��������ķ���5-10���ı仯������ATPŨ��ȴ���ֲ��䡣��ô���������ά��ATP��̬ƽ��ģ��⿴�ƼĿ�ѧ�����������о����Ǽ�ʮ�ꡣ���գ�������ѧ����ҽѧ�о������Ի����������й����ľ�ҽ��ѧ�߷���������,���֡��������ţ�mitochond...
Regulation of SRF Activity by the ATP-dependent Chromatin Remodeling Enzyme, CHD8
chromatin remodeling SRF apoptosis
2015/5/22
Under normal conditions, smooth muscle cells do not replicate, or proliferate, and provide a means of contraction for many internal organs, including blood vessels and the gut. However, under abnormal...
̽�����ݿ˵���70(heat shock protein 70��Hsp70)������Թ�����ϸ��(C2C12)��ATPˮƽ��Ӱ�졣���� ͨ����������pTRE2hyg- Hsp70�������ȶ�תȾC2C12ϸ��ϵ������Hsp70�������C2C12ϸ��ϵ���ֱ���תȾ��ϸ�������IJ�ͬʱ���(0 d��3 d��7 d)�����ϸ����ATP��ˮƽ����� Hsp70�������C2C12ϸ��ϵ��������3 d��7 d��ϸ��...
Cr(��)�Ը�ϸ��������ATPø6��ATPø8��������Ӱ���Լ���������л�ϰ��Ĺ�ϵ
�� ��ϸ�� ������ ������л ����������ø��
2013/10/29
̽��Cr(��)�Ը�ϸ��������ATPø6��ATPø8�������ˮƽ��������л��Ӱ�켰�����ϵ������ ��Cr(��) 2, 8��32 ��mol��L-1�ֱ�����������������L-02��ϸ��24 h��,��ϸ����RNA��ȡ�Լ��з���RNA,������ATPø6��ATPø8 mRNA����ˮƽ����ת¼-ӫ�ⶨ���ۺ�ø����Ӧ(qRT-PCR)�ⶨ ;ϸ��ATP�����������������������ø��������ϸ��������(ROS)������...
�۲���̴�ζ���ȱѪ/�ٹ�ע���˴�����CA1��ATPø��ϸ��ɫ������ø���Ե�Ӱ�졣 ���� :���ô�������ж�����˨���Ʊ���ȱѪ�ٹ�עģ��,��27ֻSD���Դ��������Ϊ3��:�������顢ȱѪ/�ٹ�ע�顢��̴����,�����������CA1��ATPø��COX���Եı仯�� ��� : ȱѪ/�ٹ�ע�麣��CA1��ATPø��COX���Ծ������½�(P��0.01);��̴���麣��CA1��ATPø��COX���Ը�����ȱѪ/��...
�о��ļ�ϸ��������Ƽ����ͨ����MitoKCa����������ATP���м�ͨ��(MitoKATP)��֫��Զ��Ԥ������RPC���ļ������е����á����� ����SD����72ֻ����Ϊ9�飺����ȱѪ���ࣨI/R���顢RPC�顢MitoKATP ���ż���DZ���顢MitoKCa ���ż�NS1619�顢MitoKCa ������ϼ�RPC+paxilline�顢MitoKATP ������ϼ�RPC+5-HD (5��hy...
ATP�����Լ�ͨ����U50488H����ȥ�����������յ���������ļ�ϸ���ʴ��е�����
��ͨ�� ���� ��Ƭ�� �� ��Ƭ���弤���� U50488H �ļ��� �ʺ���
2013/10/29
�о�ATP�����Լ�ͨ��(KATP)�ڦ�-��Ƭ���弤����U50488H����������ļ�ϸ���ʴ��е����á����� ��ԭ�����������������ļ�ϸ��Ϊģ�ͣ�Ӧ��ȥ����������(PE)10 ��mol��L-1�յ��ļ�ϸ���ʴ�ϸ��������Ϊ���������顢PE 10 ��mol��L-1ģ���顢5-�ǻ�����(5-HD)100 ��mol��L-1�飬���б���(Gli)50 ��mol��L-1�顢PE 10 ��mol��L-1+U504...
ϸ���ϳ�ATP��������ϵ����������������Ӵ������Լ��������ữ���̣�����Ҫ��������������ά��������������������������ATP������ͨ���ܽ���Դ��ѧ��Ӱ��ATP�ϳ���ϵ�����û��ƣ�������Դ��ѧ�����������������л���ܵĸ�ϸ��ʵ���о�ģ�ͣ�����Ԥ����Դ��ѧ�Q����Σ������Ч����ѧ��Ԥ��ʩ�ȷ������Դ��ѧ����Ÿ�ϸ��������ATP�ϳ���ϵ���о���չ������������
��������ƽ���������Ը�Ѫѹ�����ļ���������Na+,K+-ATPø,Ca2+-ATPø��Mg2+-ATPø���Ե�Ӱ��
��������ƽ ���Ը�Ѫѹ ATPø ��
2010/2/25
��������ƽ(m-Nif,ig20mg��kg-1��d-1������ҩ9��)�����������������Ը�Ѫѹ����(RVHR)Ѫѹ����������(P<0.01),�����ġ�������Na+,K+-ATPø��Ca2+-ATPø����(P<0.01),����Mg2+-ATPø����,������Ч��ϵ�о�����,m-Nif�ڽϸ���(10��1000��mol��L-1)ʱ������RVHR��������Na+,K+-ATPø��Ca2+-ATPø����,��...
�����ļ�ȱ��Ԥ������COX-2��iNOS��P38MAPK��������K+ATPͨ�����ź�ת����ϵ���о�
������ ��ͨ�� �ź�ת�� �ļ� ȱѪԤ����
2010/2/2
Ŀ�ģ�̽�ִ����ļ�ϸ��ȱ��Ԥ����(IPC)ʱ������K+ATPͨ�������ü�����P38MAPK��iNOS��COX-2�Ĺ�ϵ�� ������ ʵ���Ϊ8�飨�����顢ȱ��Ԥ�����顢IPC+HDpr�顢IPC+HDpo�顢DZX+SB�顢DZX+SMT�顢DZX+NS���DZX+HD�飩����Western blotting ���ⶨ�ļ�ϸ����COX-2��iNOS��������Phosph-P38MAPK������LDH�ͷź�...
Ŀ�ģ��۲�������ATP�����Լ�ͨ��(mitoKATP)����������ROS����ȱ���Ա����е����ü������ϵ�� ������ ������Ƭ������������ѧ������ϸ�����¼����CA1����Ⱥ����λ(PS)��ȱ��ȥ������λ(HD)�� ����� ��mitoKATP���ż�diazoxide (300 ��mol/L) Ԥ����������Ƭ�����ӳ�HD��DZ���ڼ�ȱ����PS��ʧ��ʱ�䣬��߸�����PS�Ļָ��ʡ������ÿɱ�mitoKATP...
����������ATP�����������յ�ϸ�������г�ԭͷ��ϸ���������о�
ϸ�������г� ԭͷ�� �������� ���������� ϸ������
2009/11/3
Ŀ�� ̽�ֵ������������������գ�ATP�������������յ�ϸ�������г�ԭͷ��ϸ�����������á� ���� ��������ϸ�������г�ԭͷ�ڣ��ֱ���������ɣ�5 mmol/L���顢 ATP��1.6 mmol/L���顢 �������ɣ�5 mmol/L��+ATP��1.6 mmol/L����Ϳհ����飬�����¹۲�ԭͷ�ڱ仯��ҩ���յ�8 h��ѡȡԭͷ����̬�ı������Ե�һ��Ϳհ����飬��羵�۲�������ԭͷ�ڵij��ṹ��ԭ...
Ŀ�ģ��о�ţ����Ե��������µ����������Ҽ�ϸ��Ĥ��ATP�����Լص�����IK-ATP����Ӱ�졣������ Ӧ��100%����������Һ��������ģ�ͺ�ĤƬǯȫϸ����¼����������� �������Ƚϣ��������ļ�������λʱ�̣�APD���������̣�ͬʱ�յ���IK-ATP������ţ�����ʹ����ʱ���̵�APD�ָ������������ܼ��������Ե�����IK-ATP�����ۣ� ţ������������������Ҽ�ϸ��Ĥ��ATP���м�ͨ����KATP����...
ǰ�����ض��Է��Լ����Ը�Ѫѹ�����ļ���֯ATPø���Լ�ø�ٶ���ѧ������Ӱ��
ǰ������ �ʴ� ������ �Է��Ը�Ѫѹ���� ����������ø
2009/8/24
Ŀ�ģ��۲�ǰ������(Prc-C)���Է��Ը�Ѫ ѹ����(SHR)�����Ը�Ѫѹ����(RHR)�ļ�ϸ��ĤNa+��K+ -ATPø��������Na+��K+ -ATPø��Ca2+ -ATPø��M g2+-ATPø���Լ�ø�ٶ���ѧ������Ӱ�졣���������ù���ɫ�� �ⶨATPø����,��ͨ���ⶨ��ͬATPŨ��ʱ��ATPø����,����Lineweaven-Burk��ʽ��ͼ�� ����Ȩ���Իع鷨���ֱ��,��������ø�ٷ�Ӧ����...
�й��о����������а�-��
- ���ڼ���...
�й�ѧ���ڿ����а�-��
- ���ڼ���...
�����ѧ���л������а�-��
- ���ڼ���...
�й���ѧ���а�-��
- ���ڼ���...
�ˡ���-ƪ
- ���ڼ���...
�Ρ���-ƪ
- ���ڼ���...
��������-ƪ
- ���ڼ���...
�������� -ƪ
- ���ڼ���...
֪ʶҪ��-ƪ
- ���ڼ���...
���ʶ�̬-ƪ
- ���ڼ���...
��������-ƪ
- ���ڼ���...
ѧ��ָ��-ƪ
- ���ڼ���...
ѧ��վ��-ƪ
- ���ڼ���...